
Download

Article
In vivo measurement of enhanced agouti-related peptide release in the paraventricular nucleus of the hypothalamus through Gs activation of agouti-related peptide neurons
Zhenzhong Cui1,2, Adam S. Smith3,4*
1Molecular Signaling Section, Laboratory of Bioorganic Chemistry, National Institute of Diabetes and Digestive and Kidney Diseases, Bethesda, MD 20892, USA
2Mouse Metabolism Core Laboratory, National Institute of Diabetes and Digestive and Kidney Diseases, Bethesda, MD 20892, USA
3Section on Neural Gene Expression, National Institute of Mental Health, National Institutes of Health, Bethesda, MD, USA
4Department of Pharmacology & Toxicology, School of Pharmacy, University of Kansas, Lawrence, KS, USA
2Mouse Metabolism Core Laboratory, National Institute of Diabetes and Digestive and Kidney Diseases, Bethesda, MD 20892, USA
3Section on Neural Gene Expression, National Institute of Mental Health, National Institutes of Health, Bethesda, MD, USA
4Department of Pharmacology & Toxicology, School of Pharmacy, University of Kansas, Lawrence, KS, USA
*Corresponding author: Adam S. Smith, Email: adamsmith@ku.edu
Abbreviations used: aCSF, artificial cerebrospinal fluid; AgRP, agouti-related peptide; ARC, arcuate nucleus of the hypothalamus; GCPR, G protein-coupled receptors; CNO, clozapine-N-oxide; DREADD, designer receptors exclusively activated by designer drugs; IHC, immunohistochemistry; MWCO, molecular weight cutoff; NPY, neuropeptide Y; PE, polyethylene; PVN, paraventricular nucleus of the hypothalamus
Received October 21, 2018; Revision received April 23, 2019; Accepted May 16, 2019; Published July 4, 2019
Abstract
Agouti-related peptide (AgRP) neurons of the hypothalamus play a role in hunger-triggered food intake, stability of body weight, and long-term energy balance. A recent study showed that activation of the Gs-linked G protein-coupled receptors (GCPR) expressed by hypothalamic AgRP neurons promotes a sustained increase in food intake. Enhanced AgRP release has been the postulated underlying mechanism. Here, we confirmed that activation of Gs-coupled receptors expressed by AgRP neurons in the arcuate nucleus (ARC) of the hypothalamus, which is the primary brain region for the synthesis and release of AgRP, leads to increased release of AgRP in the paraventricular nucleus of the hypothalamus (PVN). We were unable to confirm changes in AgRP expression or intracellular content using traditional histological techniques. Thus, we developed an assay to measure AgRP in the extracellular fluid in the brain using large molecular weight cut-off microdialysis probes. Our technique enables assessment of brain AgRP pharmacokinetics under physiological conditions and in response to specific pharmacological interventions designed to modulate AgRP signaling.
Keywords: AgRP, DREADD, in vivo microdialysis, arcuate nucleus of the hypothalamus
INTRODUCTION
Obesity is a burgeoning health crisis in most parts of the world [1]. It is a major risk factor for many diseases such as type 2 diabetes, cancer, and cardiovascular disease [2]. Weight loss and management are difficult to achieve via behavioral approaches, and only a limited number of appetite-suppressing drugs show modest clinical efficacy. Thus, efforts are being made to identify neuronal circuits that regulate food intake in order to more selectively target the development of new therapeutics [3,4].
There is a growing body of evidence that the neuropeptide agouti-related peptide (AgRP), which is expressed exclusively in the arcuate nucleus (ARC) of the hypothalamus, plays a critical role in controlling food intake, body weight and metabolism, and, most importantly, managing the long term stability of body weight [5-7]. For example, the firing rate of AgRP neurons is elevated in brain slices of food-deprived mice [8], and ablation of AgRP neurons in adult mice leads to starvation [9,10]. Further, the neurotransmitters AgRP and neuropeptide Y (NPY), a peptide that coexists with AgRP, increase food intake when injected into the brain [11,12]. Optogenetic and chemogenetic methods can mimic endogenous regulation of AgRP neurons [6,13-19]. Optogenetic stimulation of AgRP neurons triggers a robust increase of food intake in well-fed mice [20]. DREADDs (designer receptors exclusively activated by designer drugs) are designer G-protein coupled receptors (GPCRs) that can be activated by clozapine-N-oxide (CNO), an otherwise pharmacologically inert drug [21-24]. DREADD technology has been used to activate AgRP neurons through the Gq- and Gs-mediated signaling pathway [6,19] which mimic the activity of endogenous ghrelin and PAC1 receptors at AgRP neurons [25]. Similar to light activation, Gq or Gs DREADD stimulation of AgRP neurons leads to robust food intake, even during the light cycle when food intake is typically restricted [6,19].
It remains unclear how GPCRs signaling in AgRP neurons in the ARC affects the release of AgRP in brain regions monosynaptically connected to and down-stream from these neurons, such as the paraventricular nucleus (PVN) of the hypothalamus. This is due to the challenges of developing an assay to monitor the in vivo kinetics of AgRP, a large molecule (112 amino acids). Thus, the purpose of this study was to determine whether excitation of AgRP neurons in the ARC by DREADD technology triggers AgRP release in the PVN. Microdialysis is a sampling technique which has been extensively used for the characterization and assessment of the in vivo pharmacokinetic profile of drugs and neurotransmitters in rodents and non-human primates. Recently, with the availability of larger pore membrane probes and the push-pull method, in vivo microdialysis has been used to detect relatively large peptides in the brain (e.g., apolipoprotein E [26]; tau protein [27]; amyloid β [28]). We took advantage of this technique and investigated AgRP release in the PVN after DREADD-mediated activation of the Gs pathway in AgRP neurons. Here we report the successful monitoring of AgRP level in the mouse PVN via microdialysis.
MATERIALS AND METHODS
Animals and viruses
All experiments were conducted according to US National Institutes of Health guidelines for animal research and were approved by the NIDDK Institutional Animal Care and Use Committee. Twenty-four AgRP-ires-cre mice (Agrptm1(cre)Lowl/J; The Jackson Laboratory, stock #012899) were randomly divided into two groups and were injected bilaterally with either an adeno-associated virus encoding Gs–DREADD (AAV8-hSyn-DIO-Rs-eGFP (AV4557, titer 8 × 1012)) or a control virus (AAV8-hSyn-DIO-GFP (AV4937, titer 2 × 1012) into the ARC. Both AAVs were obtained from the UNC Vector Core Facility. Microdialysis probes were placed in the PVN during a second surgery 6 weeks after the viral injections.
Viral injections
Male mice (at least 8 weeks old) were anaesthetized with isoflurane and placed into a stereotaxic apparatus (David Kopf Instruments, model 940A with 923B mouse gas anesthesia head holder). The skull was exposed via a small incision, and holes were drilled (0.45 mm drill bits) into the skull. A 5 μl Hamilton syringe with a 33 gauge, blunt-end needle was inserted into the brain, and bilateral pressure injections of the AAV were administered targeting the ARC (Bregma: −1.46 mm AP; ± 0.3 mm ML; −5.80 mm DV). The injection volume was 200–300 nl depending on the titers of different viruses. Postoperative analgesia was provided (ketoprofen, 5 mg/kg i.p.).
In vitro microdialysis
The microdialysis probe used for in vitro study had an 8 mm shaft with a 1 mm, 1000 kDa molecular weight cutoff (MWCO) polyethylene (PE) membrane (PEP-8-01, Eicom, Kyoto, Japan). The probe was prepared by flushing it first with distilled water and then with artificial cerebrospinal fluid (aCSF) perfusion solution. The probe’s inlet and outlet were then connected to a microsyringe pump (kdScientific, KDS-250) and a peristaltic pump (LongerPump, BT100-2J with DG4-10 head), respectively, using polyethylene tubing (PE-20, 380 μm ID). Sampling was carried out at different flow rates (0.5–2.0 μl/min). Probes were placed into AgRP solutions of known concentrations (60 and 600 pg/ml), and samples were collected every 50 min and stored at −80°C until being processed.
In vivo microdialysis
Mice were anaesthetized using 2.5% tribromoethanol (Avertin®) and placed into a stereotaxic apparatus. Before the probes were implanted, a small incision was made to expose the skull, the head position was leveled using bregma and lambda as reference points, and a small hole was drilled for injections and implants. Probes were connected to the perfusion system as described for the in vitro microdialysis experiment. The pump speeds were calibrated such that the fluid perfused through the probes at a 1 μl/min flow rate. Finally, the probe was implanted in the PVN (bregma: −0.75 mm AP; 0.13 mm ML; −5.60 mm DV) and fixed using an anchoring bone screw and dental cement.
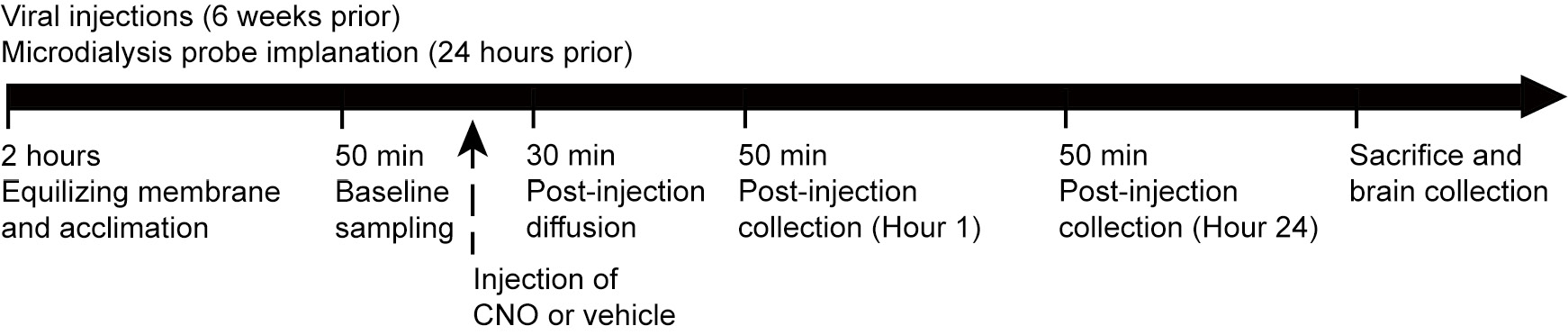
Mice were given 24 h to recover, then dialysate samples were collected every 50 min for about 2 h to establish an equilibrium between the inside and outside of the microdialysis membrane and acclimate the mice to the collection procedure (Fig. 1). Baseline samples were collected over 50 min. Mice then received a CNO injection (1 mg/kg i.p.), and the perfusion solution was allowed to flow through the probe for 30 min before the post-CNO samples were collected, at 1 and 24 h. Samples were collected into vials containing 5 µl 0.1 N HCl and immediately frozen on dry ice. All samples were stored at −80°C until processed, and all stereotaxic sites were histologically verified.
Figure 1. Graphical time-line of experimental procedure for Gs-DREADD activation of AgRP neurons and real-time in vivo microdialysis collection in mice. Eight-weeks after viral injections into the ARC and twenty-four hours after microdialysis probe implant into the PVN, mice underwent a dialysate collection protocol which included 50 min sampling at baseline and 1 and 24 h after CNO or saline administration.
AgRP ELISA
Dialysate samples from 10 mice were collected, and AgRP concentrations were measured in these samples using a mouse enzyme immunoassay kit (US Biological, 023199). However, prior to the assay, one sample from two mice, matched by condition (i.e., baseline or 1 h- or 24 h-post CNO injection), were randomly paired and pooled to yield the required sample volume and detectable concentrations for the ELISA kit. The detection limit of the enzyme immunoassay kit was 4.69 pg/ml with a sensitively of 1.83 pg/ml. The intra-assay CV was 1.44% and 3.70%, respectively.
Immunohistochemistry and imaging
To inhibit axonal transport and enhance our ability to visualize AgRP by immunohistochemical (IHC) staining, 30 µg of colchicine in 5 µl of physiological saline was delivered under anesthesia at a flow rate of 500 ng/min intracerebroventricularly (bregma: −0.34 mm AP; −1.0 mm ML; −3.0 mm DV). The mice were returned to their home cage and left undisturbed for 24 h. Then, mice were perfused with 4% paraformaldehyde in 0.1 M phosphate buffer fixative (pH 7.4), and brains were harvested and postfixed in this solution overnight. Mouse brains were sliced with a vibratome (Leica VT1000S) at 30 µm thickness and stored in 4°C in PBS until assayed. For IHC staining, brain slices were incubated overnight at 4°C with primary antibodies (either anti-AgRP, 1:1000, H-003-57, Phoenix Pharmaceuticals, INC or anti-GFP, 1:400, ab6662(FITC)) diluted in PBS, supplemented with 1% BSA and 0.1% Triton X-100. Slices were then washed three times and incubated with species appropriate and minimally cross-reactive fluorophore-conjugated secondary (Abcam; Donkey anti rabbit IgG 594, 1:000, ab150064, Abcam; goat anti rabbit 594, 1:1000) for 2 h at room temperature. Slices were rinsed in PBS with 0.1% Triton X-100 three times and mounted for imaging using Antifade Reagent (SlowFade® Gold with DAPI, S36939). Fluorescence images were taken with Zeiss and Leica fluorescent microscopes.
RNAScope method to detect AgRP expression in the ARC
Fresh frozen mouse brains were collected and stored at −80°C, then 16 µm sections were cut with a cryostat and mounted onto superfrost slides and stored at −80°C. The 2-plex 2.0 kit was used for RNAScope assays. Following the manufacturer’s (Advanced Cell Diagnostics) protocol, sections on the slides were fixed at room temperature with 10% NFB for 30 min, dried with 50%, 70% and 100% ethanol. The edges of the slides were marked with a waterproof pen. Pretreatment steps included room temperature incubations with pretreatment solution 1 (10 min), pretreatment solution 2 (10 min), and pretreatment solution 3 (30 min). Sections were hybridized at 40°C for 2 h with a specifically designed AgRP probe. Finally, sections were exposed to serial amplification steps (AMP1 through AMP6), color development, counterstaining with 10% hematoxylin, and dried by using a 65°C heat block. Cover slides were placed over the sections on the slide with Ecomount, and images were captured and processed with a Keyence Digital Microscope.
Statistical analysis
Statistical analyses were performed using IBM SPSS Statistics 19 (SPSS, Inc., an IBM Company) and were expressed as mean ± SEM. Normality was tested using Kolmogorov-Smirnov. Outliers were assessed using the IQR (interquartile range) method, denoting values three times below the 1st quartile or above the 3rd quartile as outliers. No sample calculation was performed.
RESULTS
Gs DREADD selectively expressed in ARC AgRP neurons without altering AgRP expression
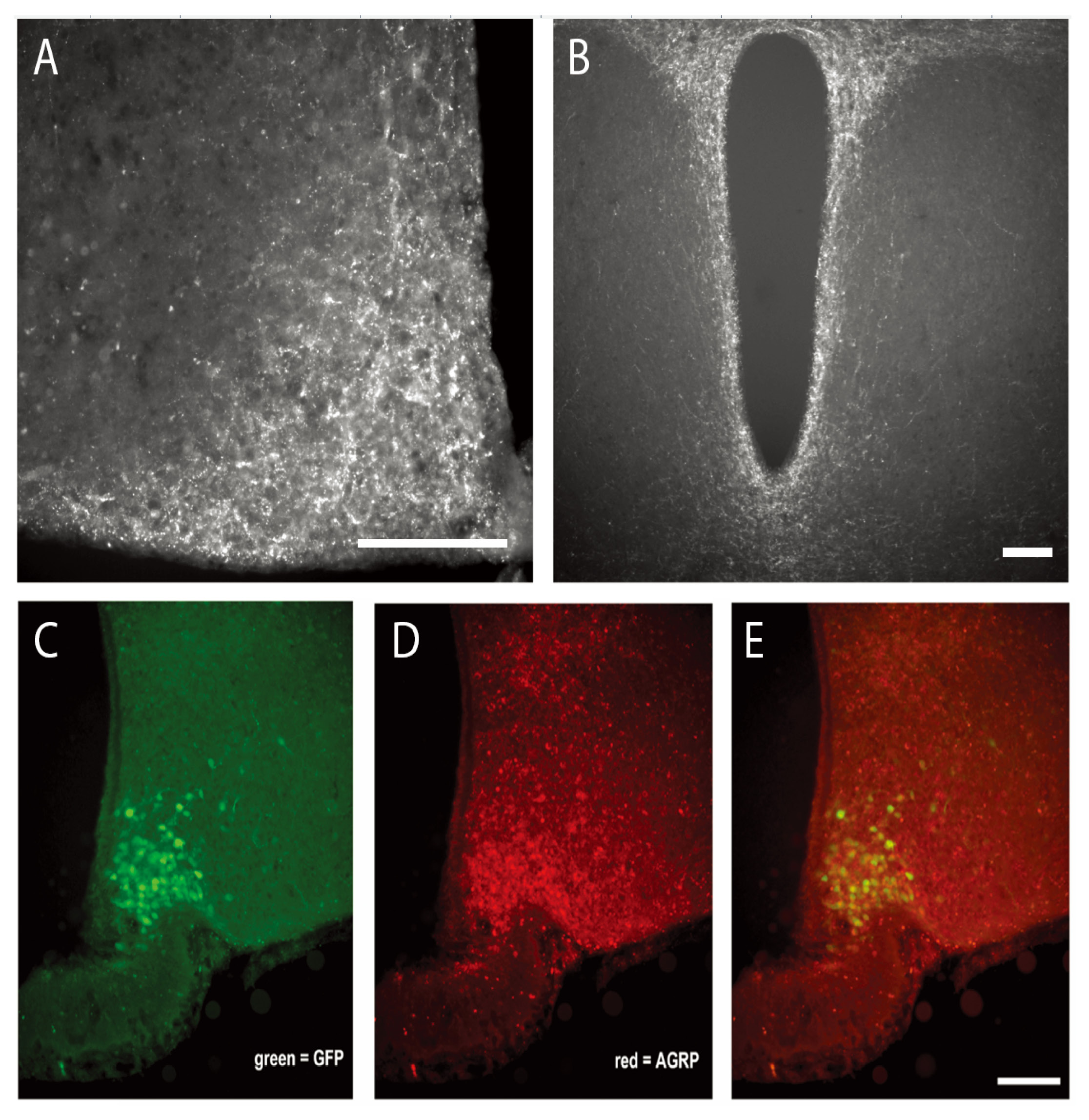
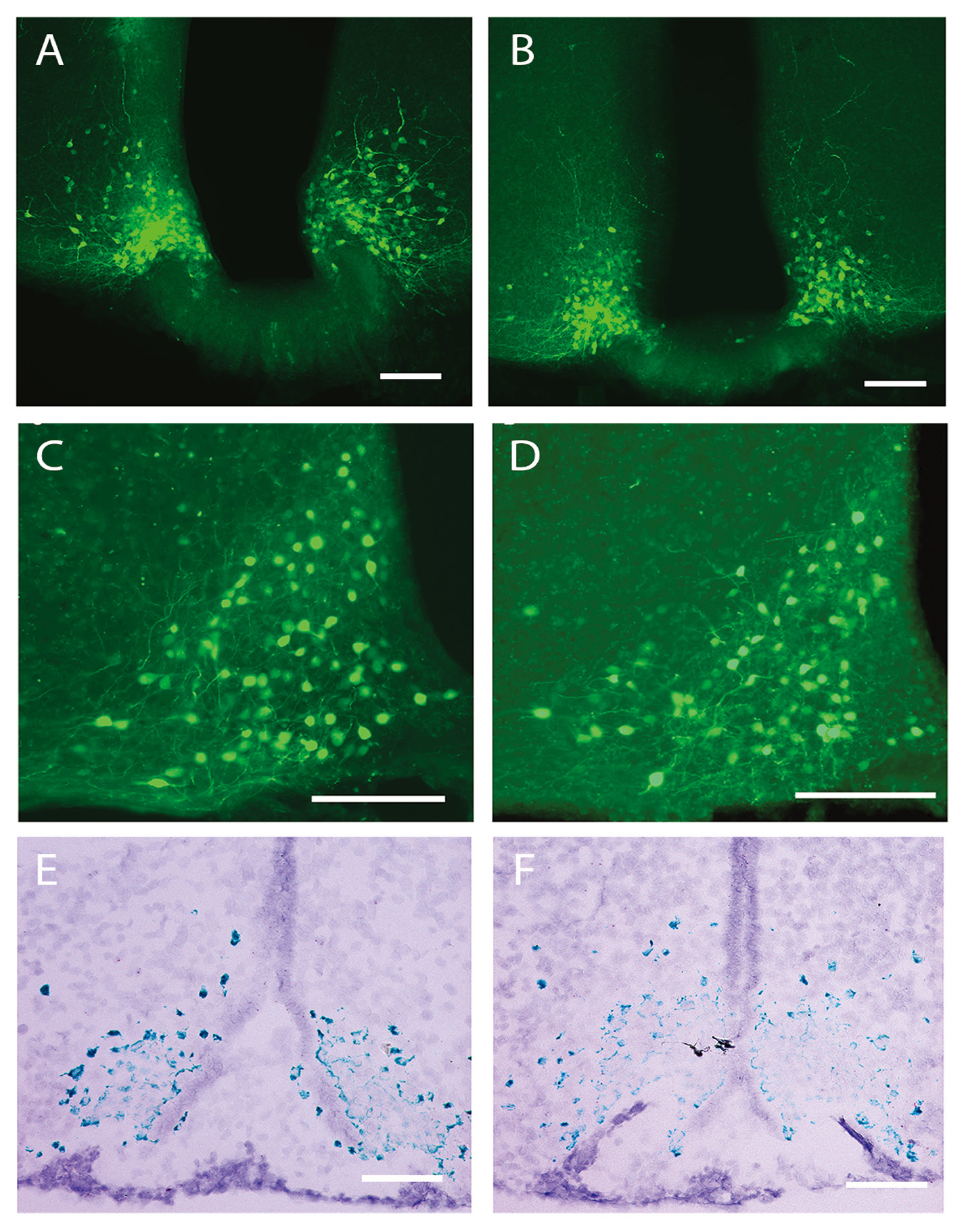
We injected Gs–DREADD or AAV-hSyn-DIO-GFP viruses into male AgRP-ires-cre mice and eight weeks later visualized the expression using IHC to visualize viral GFP expression. GFP expression was localized to cell bodies in the ARC and colocalized with AgRP expression, confirming that viral expression was selective to AgRP neurons (Fig. 2). AgRP protein and mRNA expression were quantified to determine if viral administration affected these parameters. However, neither Gs–DREADD nor AAV-hSyn-DIO-GFP administration influenced AgRP protein or mRNA levels, visualized by IHC and RNAScope, respectively (Fig. 3).
In vitro microdialysis
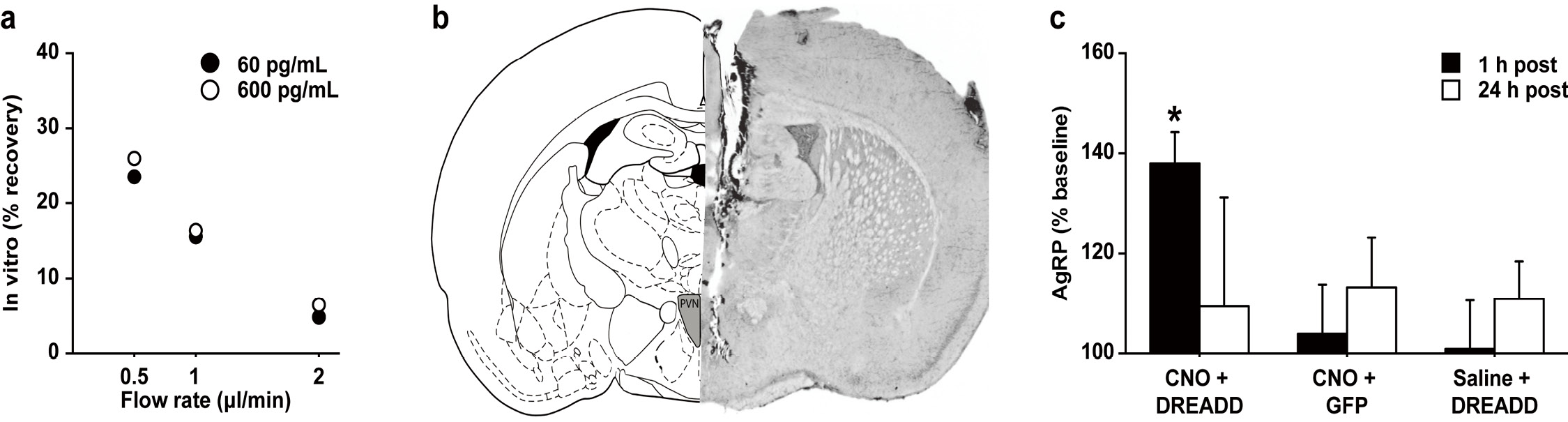
We selected a microdialysis probe membrane (1000 kDa MWCO, PEP-8-01, Eicom, Kyoto, Japan) suitable for collecting high-molecular weight molecules, as AgRP has a molecular mass of ~14 kDa. There was an inverse relationship between flow rate and percentage recovery for AgRP at a low and a high concentration, 60 and 600 pg/ml respectively (flow rate: 0.5 μl/min, 26.0% recovery; 1.0 μl/min, 17.6% recovery; 2.0 μl/min, 6.5% recovery) (Fig. 4). Based on these results, we decided to use the 1.0 μl/min flow rate for in vivo microdialysis experiments (see next paragraph).
In vivo microdialysis
To determine if Gs activation increased AgRP release, we used chemogenetic technology and real-time in vivo microdialysis to stimulate AgRP neurons in the ARC and measure subsequent AgRP release in the PVN. Specifically, we bilaterally injected the Gs-DREADD virus into the ARC of AgRP-ires-Cre mice to express the Gs-linked DREADD in AgRP ARC neurons (Fig. 2). Eight weeks later, the extracellular fluid from the PVN was collected to measure the AgRP concentrations before and after (1 h- and 24 h-post) CNO injections. Extracellular AgRP concentrations were measured 94.07 ± 2.65 pg/ml in the saline-treated mice and 90.71 ± 6.93 pg/ml in the CNO-treated mice. CNO-mediated excitation of AgRP ARC neurons markedly enhanced the release of AgRP in the PVN during the first hour (t(4) = 5.52, P < 0.005) following the CNO injection but not 24 h later (t(4) = 0.29, P = 0.79) (Fig. 4). Control mice received either the Gs-DREADD virus without CNO treatment (saline + DREADD; virus control) or were treated with CNO in the absence of Gs-DREADD expression (CNO + AAV-hSyn-DIO-GFP; drug control). No effects on AgRP release were observed for either control group.
DISCUSSION
In this study, we used in vivo microdialysis with a unique large-pore-sized membrane probe (1000-kDa MWCO) and push-pull perfusion system to monitor AgRP extracellular levels in the PVN, a major target region of hypothalamic AgRP neurons. To the best of our knowledge, this is the first report that has demonstrated the detection and characterization of AgRP release in the brain, which is important to determine the function of AgRP signaling in regulation of food intake, metabolism, and body weight.
Figure 2. Viral expression is localized to AgRP neurons in the arcuate nucleus. A. AgRP immunostaining shows AgRP expression in the arcuate nucleus following Cre-inducible AAV-hSyn-DIO-Rs-eGFP (Gs–DREADD) injection in AgRP-ires-cre mice. B. AgRP immunostaining shows AgRP expression at axon projection to paraventricular nucleus of the hypothalamus. C-E. A series of 20× images of coronal sections in the ARC of Gs–DREADD treated mice illustrating (C) GFP+, (D) AgRP expression (AgRP+, 30 µg of colchicine was injected into the lateral ventricles of mice 24 h before perfusion to enhance AgRP concentration and allow visualization of cell bodies), and (E) colocalization of both proteins. Scale bar = 100 um.
Figure 3. AgRP expression is not affected by Gs stimulation via the DREADD system. A and B. The 10× images of coronal sections show the expression of green fluorescent protein (GFP+, green) following either Cre-inducible AAV-hSyn-DIO-Rs-eGFP (Gs–DREADD) or AAV- hSyn-DIO-GFP (control virus) injection in the ARC of AgRP-ires-cre mice, respectively. C and D. Additional images of coronal sections at 40× following the same viral preparations as in panels (A) & (B). E and F. AgRP mRNA expression in the ARC after CNO treatment is similar between Gs–DREADD and control virus injected mice (A, C, E = Gs–DREADD virus; B, D, F = control virus). Scale bar = 100 um.
Figure 4. Real-time pull-push microdialysis with a large-pore-sized membrane probe proved sensitive enough to detect AgRP release in the PVN after Gs–DREADD stimulation of AgRP neurons in the ARC. A. There was an inverse relationship between flow rate and percentage recovery for AgRP at a low and high concentration in the in vitro collection of AgRP via the large-pore-sized membrane probe and push-pull microdialysis system. Each value constitutes a single sample measured. B. Schematic drawing (left) and representative photomicrograph of the mouse brain section (right) illustrate location of microdialysis probe placement in the PVN. C. Excitation of AgRP ARC neurons markedly enhanced the release of AgRP in the PVN during the first hour following the CNO injection but not 24 h later (CNO + DREAAD). Control mice were generated as either Gs-DREADD virus injected without CNO treatment (saline + DREAAD; virus control) or control virus inject with CNO treatment (CNO + GFP). No effects on AgRP release were observed for either control group. Each value constitutes 4–5 samples tested in replication and are reported as mean and standard error. * denotes a significant change from baseline (P < 0.05).
In vitro microdialysis
Microdialysis is a widely used technique to measure the release of neurotransmitters and other small molecules in the brain [29]. Low MWCO microdialysis probes (typically 6–18 kDa) prohibit collection of relatively large-sized molecules with efficiency, including peptides near the MWCO such as AgRP (~14 kDa MW). Several recent reports have demonstrated that the use of high MWCO microdialysis probes improves collection of larger molecules and peptides (e.g., apolipoprotein E [26]; tau protein [27]; amyloid β [28]). Here, we first characterized the efficiency of AgRP collection from aCSF using microdialysis in vitro. The recovery of AgRP by microdialysis was inversely related to the flow rate through the microdialysis probe, which is consistent with measurements of other analytes [29]. We determined that a 1.0 µl/min flow rate yields the best recovery rate within the largest sample volume.
In vivo microdialysis
Our in vivo microdialysis work confirmed that activation of Gs signaling in AgRP neurons activation increased release of AgRP in the PVN. Specifically, we were able to selectively express a Gs DREADD in AgRP neurons in the ARC, which was confirmed to also express in axonal fibers in the PVN. Using site-specific injections of CNO into the PVN, the AgRP concentration in the PVN was increased an hour later. PVN AgRP levels returned to baseline within 24 h after stimulation. Recent studies showed that CNO can convert in vivo into clozapine, which has binding affinity and potency to targets other than DREADD [30]. Thus, to exclude non-specific outcomes of CNO metabolite action, we administered CNO into the PVN of DREADD-free mice, which did not affect local AgRP concentrations. Previous studies indicate that AgRP affects hunger-triggered food intake, stability of body weight, and long-term energy balance [5-7], which may occur, at least in part, through signaling via Gs-linked GPCRs on AgRP neurons in the ARC [19]. There are several brain regions downstream of the ARC AgRP neurons that have garnered attention regarding food intake research, such as PVN, bed nucleus of the stria terminalis and lateral hypothalamus. The methods developed here will be useful in sampling AgRP in the extracellular fluid of these brain regions over time in order to assess the relationship between AgRP levels and feeding behaviors and the role of Gs-linked GPCRs in mediating these processes.
Several studies have shown that AgRP neuron activation alone is sufficient to trigger robust food intake [6,19,20], which has led to the characterization of these neurons as ‘hunger neurons’ [7,31,32]. Still, how precisely the activation of AgRP neurons initiates feeding is still under investigation. There is evidence that gamma-Aminobutyric acid (GABA) release from these neurons is a rapid response to inhibit downstream neurons to trigger robust feeding acutely. In contrast, Krashes and colleagues demonstrated that the AgRP-induced food intake is impaired by double inactivation of both the GABA and NPY systems, but the effect only lasted only for a few hours [31]. This suggests that AgRP release may play an important role in long term food intake. In fact, it has recently been reported that Gs pathway activation, with the same DREADD system used in the current study, promotes food intake that lasts for days and mimics the effects of i.c.v. AgRP administration [19]. In contrast, stimulation of Gq pathway with a similar DREADD system (i.e., CNO-treated hM3Dq-AgRP mice) lasted only for a single day. This may result from the difference in cellular activity caused by the activation of Gs- versus Gq-dependent signaling pathways, which may also be reflected in distinct patterns of AgRP release. The effect of Gs activation could be blocked by a selective anti-AgRP antibody. In this study, we clearly demonstrated in vivo that Gs activation increases the release of AgRP in the PVN. Thus, enhancing AgRP release or Gs signaling in AgRP neurons may represent a useful strategy to develop novel appetite-suppressing drugs.
Acknowledgments
Special thanks to Drs. Oksana Gavrilova, Hans-Jurgen Wess, and W. Scott Young as well as June Song for technical assistance with the RNAScope technique and supplying the AgRP-ires-cre mice. In addition, thanks to Drs. Gavrilova, Wess, and Young for reviewing an early version of this manuscript. This research was supported by the intramural research programs of the NIMH (ZIA-MH-002498-24) and NIDDK (DK070002).
Author contributions
Scientific concept, experimental design, and data analysis: Cui Z and Smith AS. Viral injections, immunohistochemistry, and imaging: Cui Z. In vitro and in vivo microdialysis and AgRP ELISA: Smith AS. We thank Drs. Oksana Gavrilova and Hans-Jurgen Wess for providing AgRP-ires-cre mice. We also thank Dr. W. Scott Young and June Song for performing the RNAScope to detect AgRP mRNA on tissue sections.
References
- Caballero B (2007) The global epidemic of obesity: an overview. Epidemiol Rev 29: 1-5. doi: 10.1093/epirev/mxm012. [View Article] [PubMed] [Google Scholar]
- Saper CB, Chou TC, Elmquist JK (2002) The need to feed: homeostatic and hedonic control of eating. Neuron 36: 199-211. doi: 10.1016/S0896-6273(02)00969-8. [View Article] [PubMed] [Google Scholar]
- Rueda-Clausen CF, Padwal RS, Sharma AM (2013) New pharmacological approaches for obesity management. Nat Rev Endocrinol 9: 467-478. doi: 10.1038/nrendo.2013.113. [View Article] [PubMed] [Google Scholar]
- Brownley KA, Peat CM, La Via M, Bulik CM (2015) Pharmacological approaches to the management of binge eating disorder. Drugs 75: 9-32. doi: 10.1007/s40265-014-0327-0. [View Article] [PubMed] [Google Scholar]
- Luquet S, Perez FA, Hnasko TS, Palmiter RD (2005) NPY/AgRP neurons are essential for feeding in adult mice but can be ablated in neonates. Science 310: 683-685. doi: 10.1126/science.1115524. [View Article] [PubMed] [Google Scholar]
- Krashes MJ, Koda S, Ye C, Rogan SC, Adams AC, et al. (2011) Rapid, reversible activation of AgRP neurons drives feeding behavior in mice. J Clin Invest 121: 1424-1428. doi: 10.1172/JCI46229. [View Article] [PubMed] [Google Scholar]
- Sternson SM (2013) Hypothalamic survival circuits: blueprints for purposive behaviors. Neuron 77: 810-824. doi: 10.1016/j.neuron.2013.02.018. [View Article] [PubMed] [Google Scholar]
- Baver SB, Hope K, Guyot S, Bjørbaek C, Kaczorowski C, et al. (2014) Leptin modulates the intrinsic excitability of AgRP/NPY neurons in the arcuate nucleus of the hypothalamus. J Neurosci 34: 5486-5496. doi: 10.1523/JNEUROSCI.4861-12.2014. [View Article] [PubMed] [Google Scholar]
- Wu Q, Boyle MP, Palmiter RD (2009) Loss of GABAergic signaling by AgRP neurons to the parabrachial nucleus leads to starvation. Cell 137 1225-1234. doi: 10.1016/j.cell.2009.04.022. [View Article] [PubMed] [Google Scholar]
- Wu Q, Palmiter RD (2011) GABAergic signaling by AgRP neurons prevents anorexia via a melanocortin-independent mechanism. Eur J Pharmacol 660: 21-27. doi: 10.1016/j.ejphar.2010.10.110. [View Article] [PubMed] [Google Scholar]
- Ebihara K, Ogawa Y, Katsuura G, Numata Y, Masuzaki H, et al. (1999) Involvement of agouti-related protein, an endogenous antagonist of hypothalamic melanocortin receptor, in leptin action. Diabetes 48: 2028-2033. doi: 10.2337/diabetes.48.10.2028. [View Article] [PubMed] [Google Scholar]
- Schick RR, Schusdziarra V, Nussbaumer C, Classen M (1991) Neuropeptide Y and food intake in fasted rats: effect of naloxone and site of action. Brain Res 552: 232-239. [PubMed] [Google Scholar]
- Varela L, Horvath TL (2012) Leptin and insulin pathways in POMC and AgRP neurons that modulate energy balance and glucose homeostasis. EMBO Rep 13: 1079-1086. doi: 10.1038/embor.2012.174. [View Article] [PubMed] [Google Scholar]
- Varela L, Horvath TL (2012) AgRP neurons: a switch between peripheral carbohydrate and lipid utilization. EMBO J 31: 4252-4254. doi: 10.1038/emboj.2012.287. [View Article] [PubMed] [Google Scholar]
- Sobrino Crespo C, Perianes Cachero A, Puebla Jimenez L, Barrios V, Arilla Ferreiro E (2014) Peptides and food intake. Front Endocrinol (Lausanne) 5: 58. doi: 10.3389/fendo.2014.00058. [View Article] [PubMed] [Google Scholar]
- Atasoy D, Aponte Y, Su HH, Sternson SM (2008) A FLEX switch targets Channelrhodopsin-2 to multiple cell types for imaging and long-range circuit mapping. J Neurosci 28: 7025-7030. doi: 10.1523/JNEUROSCI.1954-08.2008. [View Article] [PubMed] [Google Scholar]
- Atasoy D, Betley JN, Li W, Su HH, Sertel SM, et al. (2014) A genetically specified connectomics approach applied to long-range feeding regulatory circuits. Nat Neurosci 17: 1830-1839. doi: 10.1038/nn.3854. [View Article] [PubMed] [Google Scholar]
- Krashes MJ, Kravitz AV (2014) Optogenetic and chemogenetic insights into the food addiction hypothesis. Front Behav Neurosci 8: 57. doi: 10.3389/fnbeh.2014.00057. [View Article] [PubMed] [Google Scholar]
- Nakajima K, Cui Z, Li C, Meister J, Cui Y, et al. (2016) Gs-coupled GPCR signalling in AgRP neurons triggers sustained increase in food intake. Nat Commun 7: 10268-1038. doi: 10.1038/ncomms10268. [View Article] [PubMed] [Google Scholar]
- Aponte Y, Atasoy D, Sternson SM (2010) AGRP neurons are sufficient to orchestrate feeding behavior rapidly and without training. Nat Neurosci 14: 351-355. doi: 10.1038/nn.2739. [View Article] [PubMed] [Google Scholar]
- Adamantidis A, Arber S, Bains JS, Bamberg E, Bonci A, et al. (2015) Optogenetics: 10 years after ChR2 in neurons--views from the community. Nat Neurosci 18: 1202-1212. doi: 10.1038/nn.4106. [View Article] [PubMed] [Google Scholar]
- Guettier JM, Gautam D, Scarselli M, Ruiz de Azua I, Li JH, et al. (2009) A chemical-genetic approach to study G protein regulation of beta cell function in vivo. Proc Natl Acad Sci USA 106: 19197-19202. doi: 10.1073/pnas.0906593106. [View Article] [PubMed] [Google Scholar]
- Rogan SC, Roth BL (2011) Remote control of neuronal signaling. Pharmacol Rev 63: 291-315. doi: 10.1124/pr.110.003020. [View Article] [PubMed] [Google Scholar]
- Wess J, Nakajima K, Jain S (2013) Novel designer receptors to probe GPCR signaling and physiology. Trends Pharmacol Sci 34: 385-392. doi: 10.1016/j.tips.2013.04.006. [View Article] [PubMed] [Google Scholar]
- Cowley MA, Smith RG, Diano S, Tschop M, Pronchuk N, et al. (2003) The distribution and mechanism of action of ghrelin in the CNS demonstrates a novel hypothalamic circuit regulating energy homeostasis. Neuron 37: 649-661. doi: 10.1016/S0896-6273(03)00063-1. [View Article] [PubMed] [Google Scholar]
- Ulrich JD, Burchett JM, Restivo JL, Schuler DR, Verghese PB, et al. (2013) In vivo measurement of apolipoprotein E from the brain interstitial fluid using microdialysis. Mol Neurodegener 8: 13. doi: 10.1186/1750-1326-8-13. [View Article] [PubMed] [Google Scholar]
- Yamada K, Patel TK, Hochgräfe K, Mahan TE, Jiang H, et al. (2015) Analysis of in vivo turnover of tau in a mouse model of tauopathy. Mol Neurodegener 10: 55. doi: 10.1186/s13024-015-0052-5. [View Article] [PubMed]
- Hori Y, Takeda S, Cho H, Wegmann S, Shoup TM, et al. (2014) A Food and Drug Administration-approved asthma therapeutic agent impacts amyloid β in the brain in a transgenic model of Alzheimer disease. J Biol Chem 290: 1966-1978. doi: 10.1074/jbc.M114.586602. [View Article] [PubMed] [Google Scholar]
- Chefer VI, Thompson AC, Zapata A, Shippenberg TS (2009) Overview of brain microdialysis. Curr Protoc Neurosci Chapter 7: 1. doi: 10.1002/0471142301.ns0701s47. [View Article] [PubMed] [Google Scholar]
- Gomez JL, Bonaventura J, Lesniak W, Mathews WB, Sysa-Shah P, et al. (2017) Chemogenetics revealed: DREADD occupancy and activation via converted clozapine. Science 357: 503-507. doi: 10.1126/science.aan2475. [View Article] [PubMed] [Google Scholar]
- Sternson SM, Nicholas Betley J, Cao ZF (2013) Neural circuits and motivational processes for hunger. Curr Opin Neurobiol 23: 353-360. doi: 10.1016/j.conb.2013.04.006. [View Article] [PubMed] [Google Scholar]
- Sternson SM, Atasoy D (2014) Agouti-related protein neuron circuits that regulate appetite. Neuroendocrinology 100: 95-102. doi: 10.1159/000369072. [View Article] [PubMed] [Google Scholar]