
Download

Article
Co-culture of placental explants with isolated CD4 and CD8 T cells: a functional model to define the consequences of placental inflammation
Hayley Derricott*, Rebecca L. Jones, Alexander E.P. Heazell, Susan L. Greenwood
Maternal & Fetal Health Research Centre, University of Manchester, Manchester Academic Health Science Centre, Manchester M13 9WL, UK
*Corresponding author: Hayley Derricott, Maternal & Fetal Health Research Centre, University of Manchester, Manchester Academic Health Science Centre, Manchester M13 9WL, Phone: +44 161 7016960, E-mail: hayleyderricott@live.co.uk
Competing interests: The authors have declared that no competing interests exist.
Received August 28, 2015; Revision received November 14, 2015; Accepted December 9, 2015; Published December 22, 2015
Abstract Appropriate placental function is essential for successful pregnancy and placental dysfunction is associated with fetal growth restriction (FGR) and stillbirth. Villitis of unknown etiology (VUE) and chronic intervillositis of unknown etiology (CIUE) are immune-mediated conditions characterized by placental infiltrates of macrophages, CD4 and CD8 T cells. VUE and CIUE occur more frequently in the placentas of pregnancies complicated by FGR. The mechanisms by which this inflammation induces placental dysfunction are yet to be defined. We aimed to develop an in vitro model of placental inflammation to investigate functional consequences of immune cells in the placental environment. Fragments of placental tissue were co-cultured with CD4 and CD8 T cells isolated from whole blood. CellTrackerTM fluorescence was used to identify T cells in cultured explants. Tissue histology, endocrine and nutrient transport function was assessed using established methods. This novel preparation will enable future investigations into immune cell interactions with placenta.
Keywords: inflammation, placenta, T lymphocytes, villitis, VUE
Introduction
The placenta is critical to successful pregnancy, forming the maternal-fetal interface that supplies nutrients and oxygen to support fetal growth and removes waste products of fetal metabolism whilst maintaining a distinct barrier between mother and fetus [1]. If the placenta fails to function adequately there are serious health implications for the fetus, including an increased risk of fetal growth restriction (FGR) and stillbirth [2-5]. There is increasing evidence that sterile inflammation has a role in the placental dysfunction underlying these pregnancy complications. Villitis of unknown etiology (VUE) and chronic intervillositis of unknown etiology (CIUE) are specific inflammatory conditions of the placenta associated with FGR that may also be linked to stillbirth [6]. In the majority of cases of VUE and CIUE, an underlying clinical infection cannot be detected [7,8]. VUE is characterized by migration of CD4 and CD8 T cells from maternal blood into the stroma of the villi, the functional units of the human placenta and has been well characterized in morphological studies [9-13]. In CIUE maternal macrophages, CD4 and CD8 T cells accumulate in the intervillous space [14]. In addition to a characteristic cellular phenotype of VUE lesions, increases in pro-inflammatory cytokines interleukin (IL)-2 and IL-12 and a decrease in the anti-inflammatory cytokine IL-4 have been described in lesions [15]. Upregulation of pro-inflammatory chemokine mRNA has also been reported in VUE lesions [16]. It is probable that migrating T cells alter intravillous cytokine levels which may dysregulate placental development or function leading to increased risk of fetal harm. However, these effects have not been explored using human placenta as VUE cannot be detected antenatally. The only way to study the effects of leukocyte infiltration is by developing an in vitro model of maternal immune cell infiltration into placental tissue and examining the functional consequences.
Previous models of placental inflammation have utilized lipopolysaccharide (LPS) as a stimulator of the immune response, which mimics infectious etiology, as opposed to the sterile inflammation of VUE and CIUE [17]. Therefore, we aimed to develop an in vitro model of VUE/CIUE to enable investigations into the effect of CD4 and CD8 cell infiltration into the intervillous space and villous stroma on placental function. There is an established method of placental explant culture in which villous tissue fragments are dissected from term placental tissue and maintained in culture for 7–11 days [18]. The morphological, endocrine and functional characteristics of these cultures are well established and explants have been used extensively since 2001 to investigate the effects of long term (days) interventions on placental cell turnover, endocrinology and nutrient transport function [5,19-21]. T cells can be isolated from whole blood [22] and intracellular fluorescence can be introduced to enable cell tracking [23]. Therefore, we explored the possibility that T cells isolated from whole blood could be co-cultured with placental explants and would be visible in the intervillous space (mimicking CIUE) and in the villous stroma following migration into the tissue (mimicking the lesions in VUE).
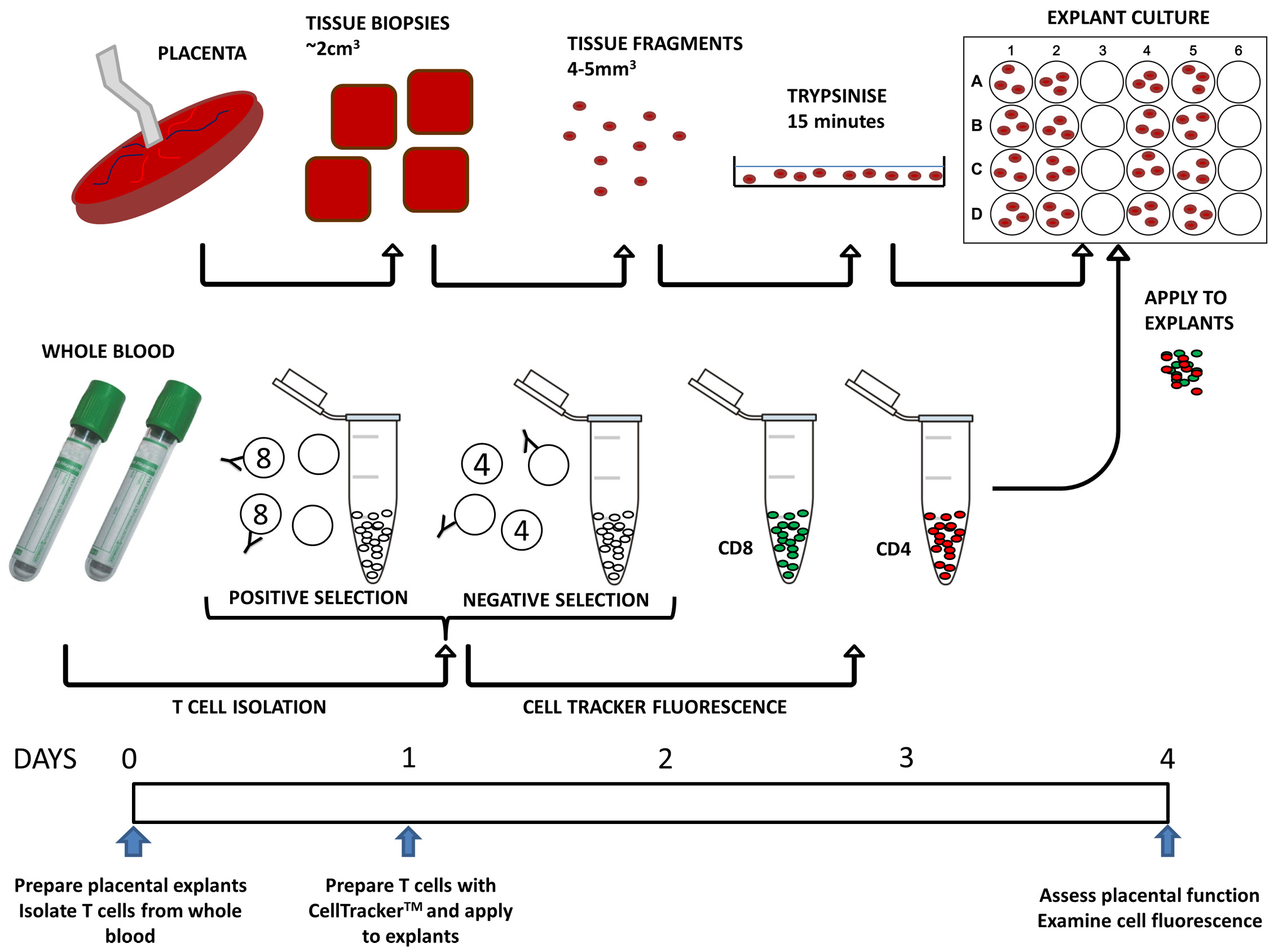
A unique feature of placental explant culture is that the syncytiotrophoblast (STB), the continuous layer of highly specialized nutrient transporting epithelium that covers the villous tissue, partially detaches in the first 1–2 days of culture and then regenerates over the course of 5–7 days to form a new STB thereafter [18]. An intact STB layer acts as a barrier to cells migrating from the intervillous space to the villous tissue. We aimed to accelerate this process in order to conform to a culture period of 4 days; the optimal time that T cells can be sustained in vitro [24]. We proposed that mild trypsinization for 15 min prior to commencing the culture would promote STB detachment without complete denudation of the underlying cytotrophoblasts and stromal tissue and encourage the passage of T cells into the tissue. A previous study that trypsinized first trimester placental tissue reported no effect on the viability of the tissue in culture [20]. We also examined whether the function of the explants was influenced by culturing them in the presence of cytokines required to maintain T cells in vitro. Endpoints were chosen to assess the effect of experimental procedures on key aspects of placental function performed by STB. Histological analysis of tissue sections stained for the cytoskeletal protein cytokeratin 7 (CK7) confirmed the integrity of STB. Endocrine function was investigated by measuring release of human chorionic gonadotrophin (hCG). This hormone is produced by differentiated STB in situ, promotes trophoblast cell fusion and stimulates progesterone production [1,25]. hCG is used as a marker of syncytialization and STB viability [26]. Its production is maximal in early pregnancy, but rises again in late pregnancy and is predominantly secreted into the maternal circulation [1]. Typically in explants, hCG release drops at day 1 of culture, gradually increasing from day 2 until a plateau is reached at around day 5 of culture [18]. Nutrient transport was assessed by measuring the activity of the system A amino acid transporter. This transporter is situated in the maternal facing membrane of STB and transfers amino acids (alanine, glycine, serine) to the fetus. System A is a well characterized sodium-dependent amino acid transporter whose activity can be determined by measuring tissue accumulation of the radiolabeled non-metabolized amino acid analogue 14Cmethylaminoisobutyric acid (MeAIB) [19,27]. System A activity in placental villous explants has been measured previously [19] and is known to be reduced in FGR [5,21,28]. The sodium-dependent uptake of 14C MeAIB into villous explants was used to determine system A activity [19]. Here we describe the results of validation experiments that have established a novel protocol for placental explant co-culture with isolated, traceable CD4 and CD8 T cells (Fig. 1).
Figure 1. Schematic representation of the timeline and steps involved in the process of explant and T cell co-culture.
Materials and Methods
Materials
All reagents supplied by Sigma-Aldrich (Dorset, UK) unless otherwise stated.
- RPMI-1640 medium (Cat. # 21875, Gibco, UK) supplemented with:
- 5% Fetal bovine serum (FBS) (Gibco, UK)
- Insulin solution 1 µg/ml
- Streptomycin sulphate 100 µg/ml
- Penicillin G 100 IU/ml
- Retinol acetate 0.1 µg/ml
- Calcium chloride
- L-alanine 25 µg/ml
- L-cysteine 200 µg/ml
- Ascorbic acid 50 µg/ml
- Sterile PBS with Ca2+ and Mg2+
- Sterile PBS without Ca2+ and Mg2+
- 70% ethanol
- 0.01% trypsin in sterile PBS
- LymphoprepTM (Cat. # 07801, Stem Cell Technologies, UK)
- EasySepTM buffer (Stem Cell Technologies, UK)
- EasySepTM CD8 T cell isolation kit, positive selection (Cat. # 18013, Stem Cell Technologies, UK)
- EasySepTM CD4 T cell enrichment kit, negative selection (Cat. # 19012, Stem Cell Technologies, UK)
- Recombinant human (rh) interleukin (IL)-2 10 ng/ml (Life Technologies, UK)
- RhIL-15 10 ng/ml (Life Technologies, UK)
- Anti-CD3 antibody 5 µg/ml (BD Pharmingen, UK)
- Anti-CD28 antibody 1 µg/ml (BD Pharmingen, UK)
- CellTrackerTM CMPTX red dye (Life Technologies, UK)
- CellTrackerTM CMFDA green dye (Life Technologies, UK)
Equipment
- Sterile Petri dishes
- 24-well culture plates
- 12-well culture plates
- Sterile Pasteur pipettes
- SepMate centrifugation tubes (Stem Cell Technologies, UK)
- 15 ml and 50 ml centrifuge tubes
- 5 ml Eppendorf tubes
- 5 ml, 10 ml and 25 ml sterile stripettes
- 5 ml round-bottomed polystyrene falcon tubes
- Sterile dissecting scissors and forceps
- Centrifuge (Beckman Coulter)
- EasySepTM magnet (Stem Cell Technologies, UK)
- Humidified incubator 37°C, 21% O2, 5% CO2
- Laminar air flow
- Inverted microscope (Olympus CK2, Olympus, Southend-on-Sea, UK)
- Fluorescent microscope (Zeiss Observer Z1 AX10, Zeiss, Welwyn Garden City, UK)
- Flow cytometer (BD Accuri C6, BD Biosciences, Oxford, UK)
Placentas and blood
Term (37–42 week) placentas were obtained within 30 min of delivery from women with uncomplicated pregnancies undergoing elective caesarean section. Tissue was donated with informed written consent in accordance with North West NHS REC approval (Ref: 08/H1010/55+5). Uncomplicated pregnancies were defined as maternal BMI < 30 and no evidence of pre-eclampsia, gestational diabetes mellitus or fetal growth restriction. Multiple pregnancies, fetal anomalies or abnormal fetal heart rate were also excluded. Blood samples used in establishing the model were taken from non-pregnant donors (10 ml) collected in lithium heparin tubes on day 0 of co-culture.
Trypsinization and syncytiotrophoblast morphology
Placental villous tissue samples were dissected, washed free of maternal blood and exposed to trypsin to accelerate STB detachment prior to culture. In initial trials, tissue was incubated in different concentrations of trypsin (0.01%–0.125%), over 5–30 min (n = 2 placentas). Following trypsin treatment, tissue was fixed in 4% neutral buffered formalin (NBF) and the effect of trypsinization on STB detachment was confirmed by examination of immunohistochemistry (IHC) to detect cytokeratin 7, a cytoskeletal protein expressed by STB. A standardized laboratory protocol for immunostaining with colorimetric detection was followed, as described by Hamilton et al. [29]. Heat-mediated antigen retrieval with citrate buffer was performed and the primary antibody applied at 0.9 µg/ml. Each tissue section had a corresponding negative control on which mouse IgG (Sigma Aldrich, Dorset, UK) was applied at the same concentration as the primary antibody.
Trypsinization and placental function
Villous tissue was maintained in co-culture with T cells for 4 days, the optimal time for maintenance of these immune cells in culture. Therefore, pilot experiments were performed to determine whether exposure to trypsin prior to culture affected STB integrity and function after 4 days compared to non-trypsinized tissue. Villous tissue was exposed to 0.01% trypsin at 37°C for 15 min, the trypsin aspirated and the tissue washed twice in RPMI 1640 medium (Gibco). Explants were cultured in a 24-well plate (3 explants per well) in RPMI 1640 medium for 96 h. Culture medium was harvested daily for hCG analysis and explants fixed daily to assess STB morphology using CK7 staining as described above. At 96 h explants were fixed in 4% neutral buffered formalin (NBF) then embedded in paraffin wax prior to immunohistochemical staining to assess apoptosis, proliferation and syncytiotrophoblast (STB) regeneration using antibodies against caspase-3, Ki67 and CK7 respectively using the method described above. Primary antibodies were applied at the following concentrations: CK7 0.9 µg/ml, Ki67 0.4 µg/ml and caspase-3 1 µg/ml. System A activity was determined as the sodium-dependent uptake of 14C MeAIB.
Trypsinization, cytokines and placental function
T cell activation, proliferation and survival, requires cytokines, specifically interleukin (IL)-2 and IL-15. This necessitates exposure of the explants to these cytokines during co-culture. To determine whether IL-2 and IL-15 alter nutrient transport, trypsinized and non-trypsinized tissue was cultured for 96 h in RPMI 1640 medium with or without IL-2 and IL-15 (both 10ng/ml) and system A activity was determined at the end of culture.
EasySep cell isolation
Initially, whole blood from a non-pregnant donor was used to evaluate the experimental protocol. The EasySep cell isolation system was used to isolate CD8 T cells by positive selection and CD4 cells by negative selection. The manufacturer’s standard protocol was followed. Briefly, whole blood was centrifuged to separate out peripheral blood mononuclear cells (PBMCs). PBMCs were then incubated with EasySep CD8 positive selection antibody before a secondary incubation with magnetic nanoparticles. The suspension was placed in a magnet to pull the CD8 cells to the side of the tube and the supernatant reserved for CD4 isolation. CD4 isolation involved incubation with an antibody cocktail to negatively select out unwanted cells and another incubation with magnetic particles. Once isolated, cells were counted and then plated at 1 × 106 cells/ml. The cells were transferred into a 12-well plate and activated via anti-CD3 coating of the wells and culture for 24 h in RPMI medium with anti-CD28 before being counted again. Control wells were uncoated and recombinant human cytokines IL-2 and IL-15 were added to half of the wells.
T cell labeling
In order to identify immune cells in the intervillous space, and to ascertain whether T cells migrated into the villous tissue, CellTrackerTM tracing dyes (Life Technologies) were identified as candidates for visualization of isolated cells. CellTrackerTM dyes are fluorescent markers that pass freely through the cell membrane becoming cell-impermeant reaction products. They are passed on to daughter cells, through 3–6 generations yet are not transferred to adjacent, non-stained cells. Isolated CD4 and CD8 T cells were cultured with CellTrackerTM Red CMPTX or CellTrackerTM Green CMFDA dyes respectively following the manufacturer’s protocol.
System A activity
To determine whether trypsinization and culture in RPMI medium with cytokines affected nutrient transport, system A activity was examined as the sodium dependent uptake of 14CMeAIB into the explants after 4 days of culture.
hCG secretion
Conditioned culture medium was collected each day. Undiluted culture medium was assayed for hCG secretion by ELISA (DRG Diagnostics, Marburg, Germany) according to standard manufacturers’ protocol. Plates were read on a FLUOstar Omega microplate reader (BMG Labtech, Aylesbury, UK), values corrected for protein content and expressed as mIU/ml/mg protein.
Procedure
Day 0 – preparation of anti-CD3 coated plates for cell culture
1.Prepare 4 ml sterile PBS supplemented with 5 µg/ml anti-CD3 antibody.
2.Add 1 ml antibody solution to 4 wells of a 12-well plate.
3.Incubate for 2 h at 37°C.
4.Wash twice with 1 ml sterile PBS.
NOTE: Coated plates can be prepared the day before they are required and stored at 4°C overnight instead of a 2-h incubation at 37°C on the day of isolation.
Day 0 – production of placental fragments for culture
5.Prepare 1 L RPMI-1640 medium supplemented with 5% fetal bovine serum (FBS), 1 µg/ml insulin, 100 µg/ml streptomycin sulphate, 100 IU/ml penicillin G, 0.1 µg/ml retinol acetate, 25 µg/ml L-alanine, 200 µg/ml L-cysteine and 50 µg/ml ascorbic acid. Filter prepared medium through 0.2 µm bottle top vacuum filter (Corning, Germany).
6.Receive placenta within 30 min of delivery. Take four biopsies of approximately 1 cm3 from random areas of parenchyma, place in pre-warmed sterile PBS. All subsequent steps take place in the laminar flow hood.
7.Cut approximately 30 smaller fragments (~4–5 mm3) of villous tissue from each of the large biopsies, taking care to avoid fetal chorionic plate, decidua and large blood vessels. Transfer to sterile petri dishes with 10 ml pre-warmed sterile PBS.
8.Wash twice in sterile PBS.
9.Transfer tissue fragments to an empty petri dish. Add 10 ml 0.01% trypsin, incubate at 37°C for 15 min.
10.Remove trypsin solution then neutralize with two changes of RPMI medium.
11.Add 1 ml RPMI medium to 16 wells of a 24 well culture dish and place three fragments in each
12.Culture for 24 h in a humidified incubator at 37°C, 21% O2.
Day 0 – T cell isolation from 10 ml whole blood
13.Add 15 ml Lymphoprep to a SepMate tube.
14.In a separate centrifuge tube, mix blood with an equal volume of EasySep buffer. Carefully transfer the blood/buffer mix to the SepMate tube ensuring the tube is kept vertical.
15.Centrifuge at 1200 g for 10 min at room temperature with the brake on.
16.Pour off the supernatant (which contains the peripheral blood mononuclear cells) into a new centrifuge tube; do not invert the SepMate tube for longer than two seconds. NOTE the tube that the supernatant is poured into does not have to be a SepMate tube.
17.Centrifuge for 8 min at 300 g at RT.
18.Remove supernatant with a stripette taking care not to disturb the pellet.
19.Add ~10 ml EasySep buffer and re-suspend the cells. Centrifuge for 8 min at 300 g at RT.
20.Repeat steps 6 and 7.
21.Remove supernatant and re-suspend cells in 4 ml EasySep buffer. Pour into a 5 ml round-bottomed falcon tube.
22.Centrifuge for 8 min at 300 g at RT, remove supernatant.
23.Re-suspend pellet with 100 µl EasySep buffer. Add 10 µl EasySep CD8 positive selection cocktail. Mix well and incubate for 15 min at RT.
24.Mix the magnetic nanoparticles by vigorous pipetting (do not vortex). Add 10 µl of magnetic nanoparticles. Mix well, incubate for 10 min at RT.
25.Bring the cell suspension to a total of 2500 µl using EasySep buffer. Mix by gentle pipetting. Place the tube in the magnet for 5 min ensuring that tube does not have the cap on.
26.Whilst the tube is still in the magnet invert it, pouring the supernatant into a new 5 ml falcon tube. Leave inverted for 2–3 seconds but do not shake or blot any drops. Save the pour off for the isolation of CD4 T cells.
27.Take the tube out of the magnet, add ~2500 µl EasySep buffer. Mix by gentle pipetting, place the tube back in the magnet and set aside for 5 min.
28.Whilst the tube is still in the magnet invert it, pouring off the supernatant. (There is no need to save this supernatant.)
29.Repeat steps 15 and 16.
30.Re-suspend in 2500 µl EasySep buffer, centrifuge for 8 min at 300 g at RT.
31.Remove supernatant, re-suspend in 2 ml RPMI medium with 1 µg/ml anti-CD28, 10 ng/ml rhIL-2 and 10 ng/ml rhIL-12.
32.Use the supernatant/first pour off from the CD8 selection, centrifuge for 8 min at 300 g at RT.
33.Remove the supernatant, re-suspend cells in 250 µl EasySep buffer.
34.Add 12.5 µl EasySep Human CD4 T cell enrichment cocktail. Mix well and incubate for 10 min at RT.
35.Vortex the D magnetic particles for 30 seconds. Add 25 µl magnetic particles, mix well and incubate for 5 min at RT.
36.Bring the cell suspension to 2500 µl by adding EasySep buffer (~2200 µl) and mix gently.
37.Place the tube, without cap, into the magnet for 5 min.
38.With the tube in the magnet, invert, pouring the supernatant into a new 5 ml tube. The cells are in the pour off.
39.Centrifuge for 8 min at 300 g at RT.
40.Remove supernatant, re-suspend in 2 ml RPMI medium with 1 µg/ml anti-CD28 to activate T cells, 10 ng/ml rhIL-2 and 10 ng/ml rhIL-15 to stimulate proliferation of T cells.
41.Plate cells into the coated wells of a 12-well plate.
42.Incubate overnight in a humidified incubator at 37°C, 21% O2.
Day 1 – Introduction of CellTrackerTM fluorescence
43.Remove medium from wells, transferring into two 5 ml Eppendorfs (one for CD4 cells, one for CD8 cells).
44.Add 500 µl ice-cold sterile PBS (without Ca2+ and Mg2+), gently scrape the bottom of the wells to release the cells. Transfer to the Eppendorfs containing the medium.
45.Examine the plates on an inverted microscope to check how many cells remain attached.
46.Repeat step 2.
47.Centrifuge at 1300 rpm for 5 min at RT.
48.Dilute 1 µl CellTracker red in 1 ml pre-warmed RPMI medium and 1 µl CellTracker green in 1 ml pre-warmed RPMI medium.
49.After centrifugation, remove supernatant, re-suspend cells in pre-warmed medium with CellTracker (we used red for CD4 cells and green for CD8 cells). Incubate at 37°C for 45 min.
50.Centrifuge at 1300 rpm for 5 min at RT.
51.Re-suspend in 1 ml pre-warmed medium. Incubate at 37°C for 30 min.
52.Centrifuge at 1300 rpm for 5 min at RT.
53.Re-suspend in 1 ml pre-warmed sterile PBS (with Ca2+ and Mg2+). Centrifuge at 1300 rpm for 5 min at RT.
54.Re-suspend in 480 µl medium supplemented with rhIL-2 and rhIL-15.
Day 1 – Application of cell suspension to placental explants
55.Collect medium from 2 wells for analysis.
56.Aspirate medium from other wells.
57.Add 10 µl cell suspension (~16,000 cells (range 4,000-42,000) to each explant.
58.Incubate for 24 h in a humidified incubator at 37°C, 21% O2.
Day 2 – Add medium
59.Add 300 µl RPMI medium with rhIL-2 and rhIL-15 to each well.
60.Incubate for 24 h in a humidified incubator at 37°C, 21% O2.
Day 3 – Change medium
61.Collect medium from 2 wells for analysis.
62.Aspirate medium from other wells.
63.Add 1 ml RPMI medium with rhIL-2 and rhIL-15 to each well.
64.Incubate for 24 h at 37°C, 21% O2, 95% humidity.
Day 4 – Harvest explants and process for assessment of placental function
65.Collect medium from two wells for analysis.
Results
Trypsinization and syncytiotrophoblast morphology
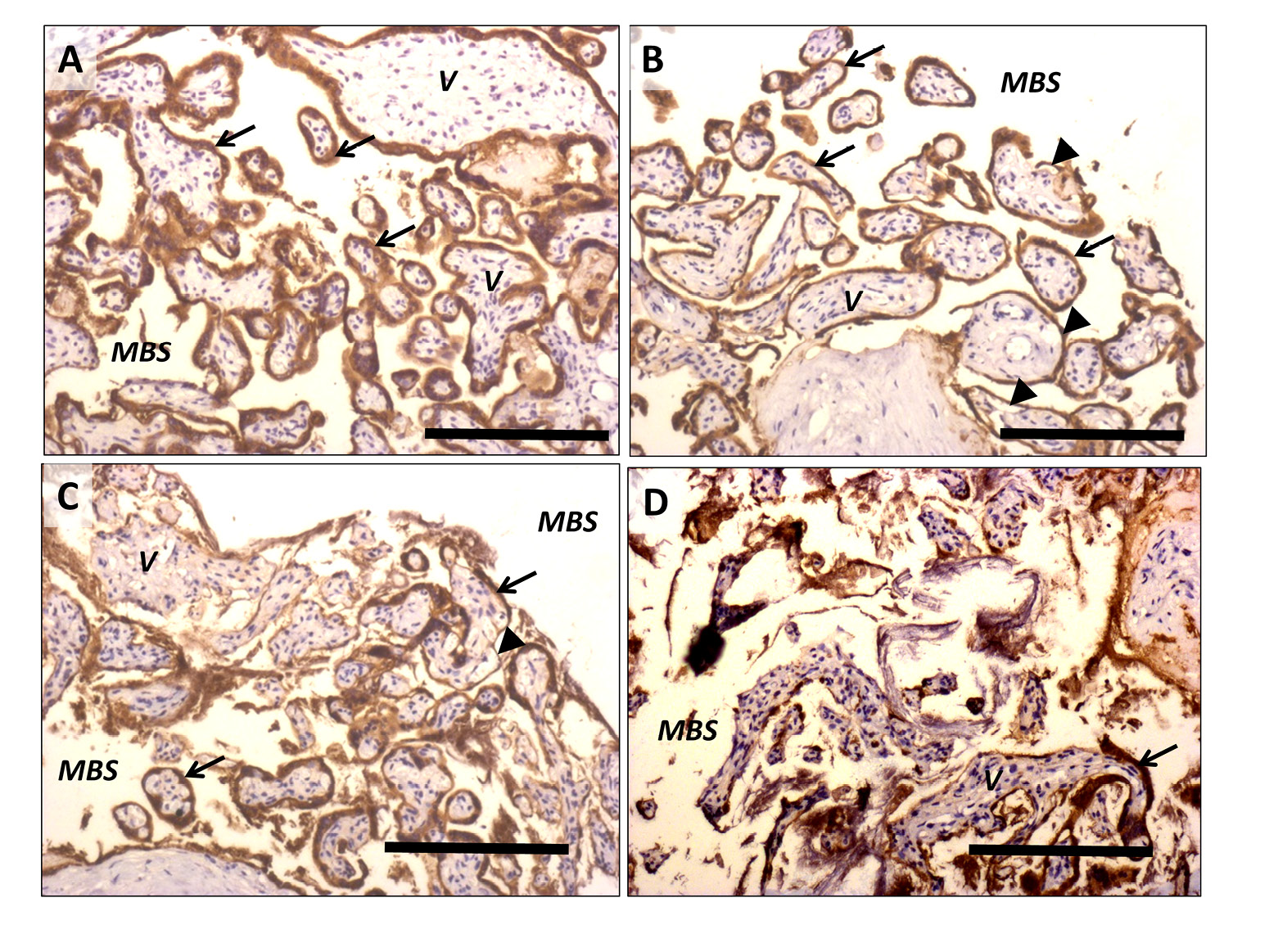
A concentration of 0.01% trypsin for 15 min was found to be sufficient to accelerate the shedding process without affecting the integrity of the villous stroma (Fig. 2).
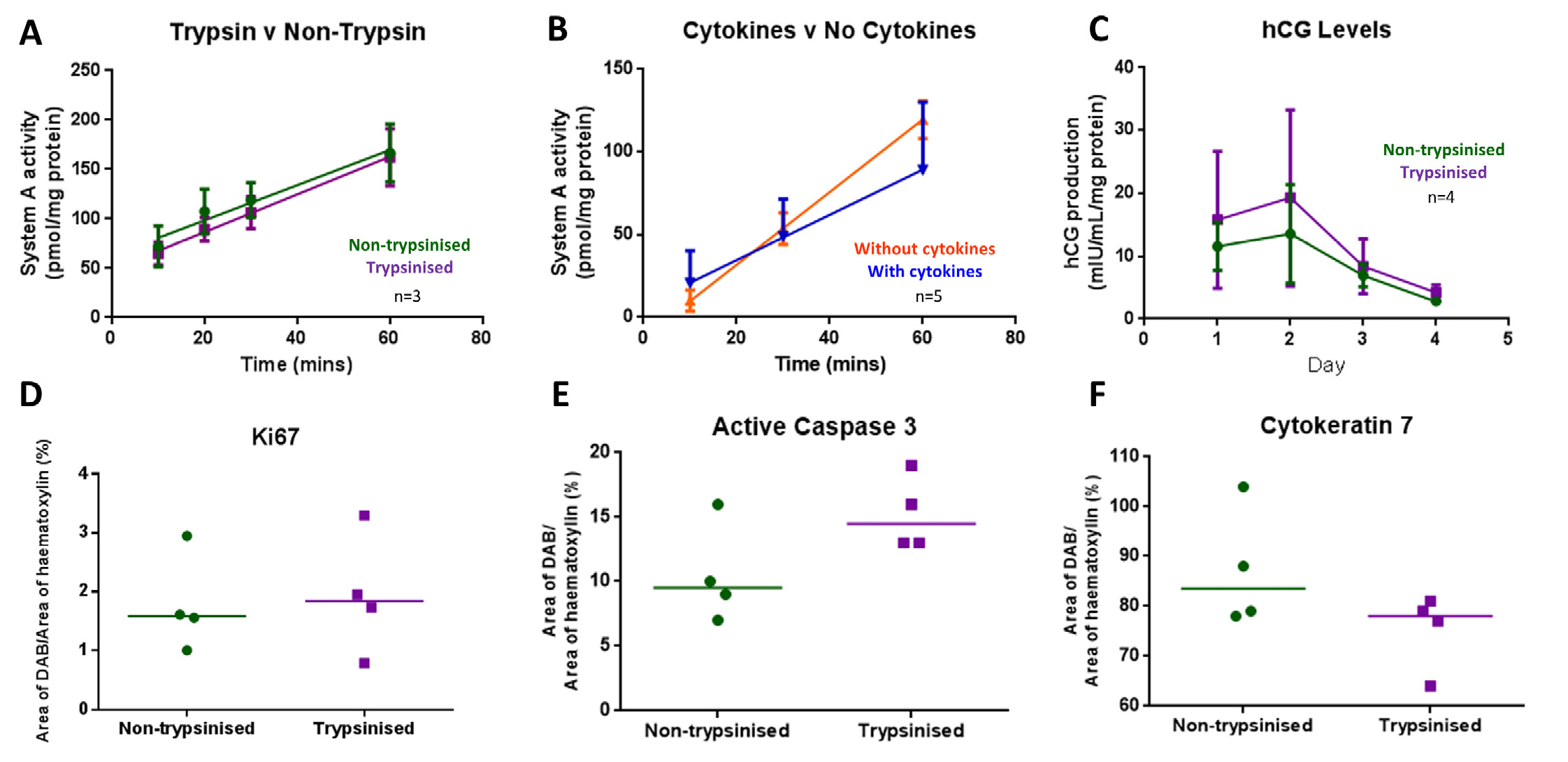
Qualitative analysis of CK7 staining from the 96 h tissue demonstrated that the morphology of the explants exposed to trypsin prior to culture was comparable to the non-trypsinized (Fig. 3F). Additionally, System A activity was not considered to be irreparably affected by trypsinization.
T cell labeling
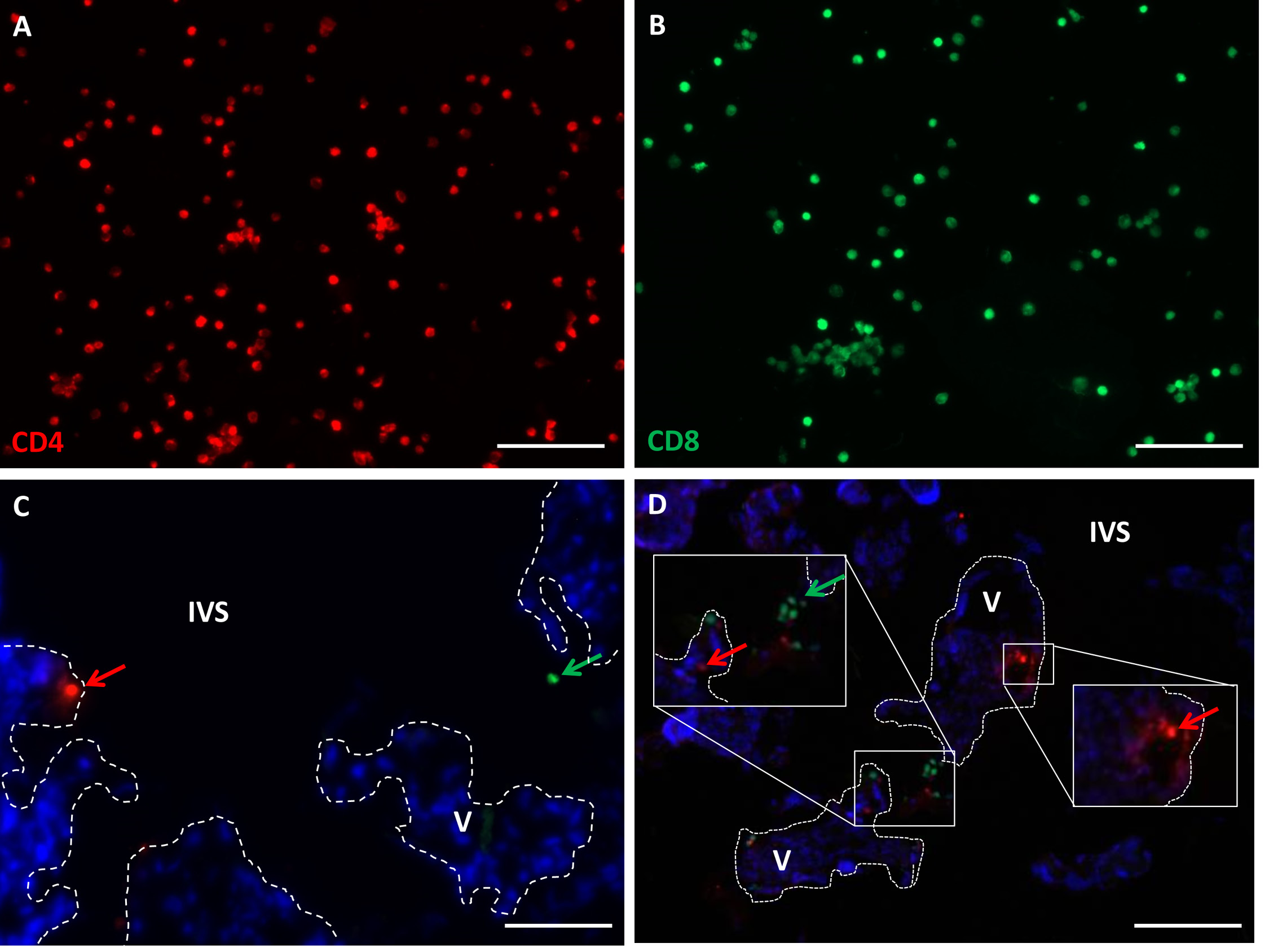
Figure 4 illustrates that the fluorescent dyes are adequately incorporated into the isolated T cells.
System A activity
System A activity was unaffected by culture in IL-2 and IL-15 supplemented medium. Figure 3 shows that system A activity over a 60 min period remained linear and was not affected by trypsinization (n = 3) or culture in medium containing cytokines (n = 5) compared to untreated explants (controls).
Hormone secretion
Endocrine function of the STB was determined by measuring hCG secretion into culture medium. hCG secretion was not affected by pre-culture exposure of tissue fragments to trypsin (n = 4, Fig. 3C) or by the addition of cytokines to culture medium (n = 2, data not shown).
Proliferation and apoptosis
Cell proliferation and apoptosis in explants after 4 days of culture (n = 3) was assessed using IHC with antibodies against Ki67 (proliferation) and active caspase-3 (apoptosis) as previously described [30]. HistoQuest (TissueGnostics, Austria), unbiased image analysis software, was used to quantify DAB staining. DAB+ areas or events were expressed as a percentage of total haematoxylin+ areas or events. There was no significant difference in levels of proliferation or apoptosis in the trypsinized explants compared to controls (Fig. 3D and E) or the explants cultured in medium supplemented with cytokines compared to those cultured in medium alone (data not shown).
Isolated T cells
Flow cytometric analysis of the CD4 and CD8 T cells demonstrated that ~80-90% purity had been achieved. After 24 h in culture an average cell yield of 1.7 × 106 CD4 T cells (range 5.1 × 105–4.1 × 106) and 1.5 × 106 CD8 T cells (4.2 × 105–3.8 × 106) was achieved. It is normal to encounter different proportions of CD4 and CD8 T cells between individuals. In our samples CD4 T cells accounted for 85-95% of those isolated and CD8 T cells 5–15%.
Isolated T cells in explants
In further pilot studies, fluorescence imaging was employed to ascertain whether isolated CD4 and CD8 T cells could be located in the vicinity of villous tissue. This confirmed that Cell Tracker persisted for the 3 days that the cells were required to be in co-culture and localized maternal cells to the areas of the placenta associated with VUE. Figures 4C and D show representative images of placental explants with fluorescing CD4 (red) and CD8 (green) T cells in the explant. Qualitative analysis of the explants that were co-cultured with T cells showed evidence of sparse cell infiltration; T cell presence was not observed in every field of view. Trypsinized explants presented areas of tissue with slightly more extensive T cell presence (Fig. 4C and D).
Figure 2. Representative images of placental tissue fragments stained with cytokeratin 7 following incubation in trypsin (0.01–0.125%) for 5-30 min. When intact, syncytiotrophoblast staining positive for cytokeratin 7 can be observed adherent to the villous tissue. Arrows show areas where the syncytiotrophoblast and villous stroma remain completely intact, arrowheads show areas where the syncytiotrophoblast layer is intact but has started to lift from the villous stroma A) Fragments incubated with 0.01% trypsin for 5 min. B) 0.01% trypsin for 15 min (the concentration used in the final protocol). C) 0.05% trypsin for 15 min. There are areas where syncytiotrophoblast has completely come away from the villous tissue and has started to disintegrate. D) 0.125% trypsin for 30 min. Large areas of complete denudation, degeneration and vacuolation of syncytiotrophoblast and villous stroma are visible. V: villous stroma, MBS; maternal blood space. Scale bars represent 100 µm, original magnification ×100.
Figure 3. Assessment of placental function following pre-culture trypsinization and culture with cytokines interleukin (IL)-2 and IL-15 at day 4 of culture. A. System A activity, measured as the sodium-dependent tissue accumulation of the radiolabeled substrate 14CMeAIB over time, was unaffected by pre-culture exposure to trypsin (mean±SE of 3 placentas), B. System A activity was unaffected by the presence of cytokines in the culture medium (mean±SE of 5 placentas), C. hCG secretion was not altered by pre-culture exposure to trypsin (mean±SE of 4 placentas). D-F. There was no significant difference in immunostaining for proliferation (Ki67) (D), apoptosis (caspase-3) (E), STB regeneration (cytokeratin 7) (F). All expressed as DAB+ area as a percentage of haematoxylin+ area between trypsinized and non-trypsinized explants.
Figure 4. Representative images of isolated CD4 and CD8 T cells. A. CD4 T cells fluorescing red after the addition of Cell Tracker CMPTX. B. CD8 T cells fluorescing green after the addition of Cell Tracker CMFDA green. C and D. CD4 T cells (red arrows) and CD8 T cells (green arrows) in placental explant tissue and intervillous space at day 4 of co-culture. Nuclei are stained with DAPI. V: villous stroma, IVS: intervillous space. Dotted lines demarcate STB. Scale bars represent 20 µm, original magnification ×200.
Discussion
The developmental work presented here has shown that it is possible to co-culture placental explants and T cells isolated from whole blood. The pilot data confirm that exposure of placental tissue to mild trypsinization prior to culture does not compromise the functional capacity of the tissue. This was shown after examination of nutrient transport function via system A activity, endocrine secretion (hCG) or cellular kinetics (proliferation and apoptosis). In addition, the cytokines required to activate and stimulate proliferation of T cells (IL-2 and IL-15) do not adversely affect placental function. CD4 and CD8 T cells can be isolated, purified and fluorescently labeled, and their viability can be maintained for 4 days in culture. They can also be observed infiltrating the intervillous space and stroma of pre-trypsinized placental villous tissue over this time frame.
For these validation experiments, blood from non-pregnant donors was used to ascertain protocol feasibility. However, in order to mimic VUE conditions the protocol would require the isolation of maternal T cells from blood. Maternal blood obtained at the time of delivery could be utilized, for example obtaining samples of maternal blood and placenta at elective Caesarean section; the timing of the protocol allows for this.
VUE is described as an inflammatory condition characterized by a villous infiltrate of fetal macrophages (Hofbauer cells; HBC) and maternal T cells [7,11,16,31]. It has been suggested that HBCs may interact with infiltrating maternal cells and thus, play a role in VUE [31]. We have previously shown that HBC numbers are increased in pregnancy pathologies such as FGR and stillbirth [15]. While individual variation in HBC number between placentas may influence the in vitro interaction or infiltration of T cells with placental tissue, this scenario is likely to occur in vivo as well. Critically, as there is no diagnostic test for VUE prior to birth, there is no means by which VUE, CIUE or numbers of HBCs can be evaluated in a sufficiently timely manner to allow functional experiments to be conducted on placental samples. An alternative approach would be to conduct functional experiments on unselected placental samples and subsequently to analyze their results by VUE status after a diagnosis has been made. However, as the incidence of VUE at term is estimated at 12%, 8-fold more experiments would be required, thus such an approach is unrealistic. Both these observations support the need for the development of in vitro models of placental inflammation to understand its functional consequences.
Our novel experimental data provides a working model on which to base co-culture studies to explore the mechanisms by which T cell infiltration could cause placental dysfunction in VUE. However, the use of this model could be advocated for a broader spectrum of experimental conditions including CIUE, diabetes-induced inflammation and autoimmune conditions such as systemic lupus erythematosus. The role of a range of specific leukocytes in inflammatory conditions could be examined as could the mechanism by which tolerance of a pregnancy is achieved.
Troubleshooting
The preliminary studies carried out to design this protocol used blood from non-pregnant donors. For experiments to examine the mechanics of VUE, placenta and maternal blood from the same pregnancy would be used. On day 0 if the placenta and blood samples arrive at the laboratory at the same time, processing of the placenta should take priority as its functional capacity depletes in a shorter time frame.
If Cell Tracker cytoplasmic fluorescence fails, ensure that the vials have been protected from light during storage.
Acknowledgments
This work was completed by HD as part of a PhD project funded by Tommy's - the Baby Charity.
References
- Gude NM, Roberts CT, Kalionis B, King RG (2004) Growth and function of the normal human placenta. Thromb Res 114: 397-407. doi: 10.1016/j.thromres.2004.06.038. [View Article] [PubMed] [Google Scholar]
- Miller J, Turan S, Baschat AA (2008) Fetal growth restriction. Semin Perinatol 32: 274-280. doi: 10.1053/j.semperi.2008.04.010. [View Article] [PubMed] [Google Scholar]
- Mullins E, Prior T, Roberts I, Kumar S (2012) Changes in the maternal cytokine profile in pregnancies complicated by fetal growth restriction. Am J Reprod Immunol 68: 1-7. doi: 10.1111/j.1600-0897.2012.01132.x. [View Article] [PubMed] [Google Scholar]
- Scifres CM, Nelson DM (2009) Intrauterine growth restriction, human placental development and trophoblast cell death. J Physiol 587: 3453-3458. doi: 10.1113/jphysiol.2009.173252. [View Article] [PubMed] [Google Scholar]
- Desforges M, Sibley CP (2010) Placental nutrient supply and fetal growth. Int J Dev Biol 54: 377-390. doi: 10.1387/ijdb.082765md. [View Article] [PubMed] [Google Scholar]
- Derricott H, Jones RL, Heazell AEP (2013) Investigating the association of villitis of unknown etiology with stillbirth and fetal growth restriction - a systematic review. Placenta 34: 856-862. doi: 10.1016/j.placenta.2013.07.003. [View Article] [PubMed] [Google Scholar]
- Redline RW, Patterson P (1993) Villitis of unknown etiology is associated with major infiltration of fetal tissue by maternal inflammatory cells. Am J Pathol 143: 473-479. [PubMed] [Google Scholar]
- Feeley L, Mooney EE (2010) Villitis of unknown aetiology: correlation of recurrence with clinical outcome. J Obstet Gynaecol (Lahore 30: 476-479. doi: 10.3109/01443611003802339. [View Article] [PubMed] [Google Scholar]
- Brito H, Juliano P, Altemani C, Altemani A (2005) Is the immunohistochemical study of the inflammatory infiltrate helpful in distinguishing villitis of unknown etiology from non-specific infection villitis?. Placenta 26: 839-841. doi: 10.1016/j.placenta.2004.10.012. [View Article] [PubMed] [Google Scholar]
- Benzon S, Zekić Tomaš S, , Benzon Z, Vulić M, Kuzmić Prusac I (2015) Involvement Of T Lymphocytes In The Placentae With Villitis Of Unknown Etiology From Pregnancies Complicated With Preeclampsia. J Matern Neonatal Med Mar 27: 1-6. doi: 10.3109/14767058.2015.1032239. [View Article] [PubMed] [Google Scholar]
- Myerson D, Parkin RK, Benirschke K, Tschetter CN, Hyde SR (2006) The pathogenesis of villitis of unknown etiology: analysis with a new conjoint immunohistochemistry-in situ hybridization procedure to identify specific maternal and fetal cells. Pediatr Dev Pathol 9: 257-265. doi: 10.2350/08-05-0103.1. [View Article] [PubMed] [Google Scholar]
- Labarrere CA, Hardin JW, Haas DM, Kassab GS (2015) Chronic villitis of unknown etiology and massive chronic intervillositis have similar immune cell composition. Placenta 36: 681-686. doi: 10.1016/j.placenta.2015.03.008. [View Article] [PubMed] [Google Scholar]
- Derricott H, Jones RL, Batra G, Heazell AEP (2014) Characterising Villitis of Unknown Etiology in Stillbirth. Reprod Sci 21: 179A.[Google Scholar]
- Contro E, deSouza R, Bhide A (2010) Chronic intervillositis of the placenta: a systematic review. Placenta 31: 1106-1110. doi: 10.1016/j.placenta.2010.10.005. [View Article] [PubMed] [Google Scholar]
- Derricott H, Jones RL, Batra G, Greenwood SL (2015) Cell and Cytokine Profile of VUE Lesions in Stillbirth. Reprod Sci 22: 378A–379A.[Google Scholar]
- Kim MJ, Romero R, Kim CJ, Tarca AL, Chhauy S, et al. (2009) Villitis of unknown etiology is associated with a distinct pattern of chemokine up-regulation in the feto-maternal and placental compartments: implications for conjoint maternal allograft rejection and maternal anti-fetal graft-versus-host disease. J Immunol 182: 3919-3927. doi: 10.4049/jimmunol.0803834. [View Article] [PubMed] [Google Scholar]
- Xiao J, Garcia-Lloret M, Winkler-Lowen B, Miller R, Simpson K, et al. (1997) ICAM-1-Mediated Adhesion of Peripheral Blood Monocytes to the Maternal Surface of Placental Syncytiotrophoblasts. Am J Pathol 150: 1845-1860. [PubMed] [Google Scholar]
- Simán CM, Sibley CP, Jones CJ, Turner MA, Greenwood SL (2001) The functional regeneration of syncytiotrophoblast in cultured explants of term placenta. Am J Physiol Regul Integr Comp Physiol 280: [PubMed] [Google Scholar]
- Audette MC, Greenwood SL, Sibley CP, Jones CJP, Challis JRG, et al. (2009) Dexamethasone stimulates placental system A transport and trophoblast differentiation in term villous explants. Placenta 31: 97-105. doi: 10.1016/j.placenta.2009.11.016. [View Article] [PubMed] [Google Scholar]
- Forbes K, Westwood M, Baker PN, Aplin JD (2008) Insulin-like growth factor I and II regulate the life cycle of trophoblast in the developing human placenta. Am J Physiol Cell Physiol 294: doi: 10.1152/ajpcell.00035.2008. [View Article] [PubMed] [Google Scholar]
- Lager S, Powell TL (2012) Regulation of nutrient transport across the placenta. J Pregnancy 2012: 179827. doi: 10.1155/2012/179827. [View Article] [PubMed] [Google Scholar]
- Chen Q, Kwang J, He F, Chan JKY, Chen J (2012) GM-CSF and IL-4 Stimulate Antibody Responses in Humanized Mice by Promoting T, B and Dendritic Cell Maturation. J Immunol 189: 5223-5229. doi: 10.4049/jimmunol.1201789. [View Article] [PubMed] [Google Scholar]
- Grolleau-Julius A, Harning EK, Abernathy LM, Yung RL (2008) Impaired Dendritic Cell Function in Aging Leads to Defective Antitumor Immunity. Cancer Res. Cancer Res 2008: 6341-6349. doi: 10.1158/0008-5472.CAN-07-5769. [View Article] [PubMed] [Google Scholar]
- Jelley-Gibbs DM, Lepak NM, Yen M, Swain SL (2000) Two distinct stages in the transition from naive CD4 T cells to effectors, early antigen-dependent and late cytokine-driven expansion and differentiation. J Immunol 165: 5017-5026. [PubMed] [Google Scholar]
- Evain-Brion D, Malassine A (2003) Human placenta as an endocrine organ. Growth Horm IGF Res 13: 34-37.[Google Scholar]
- Omata W, Ackerman 4th WE, Vandre DD, Robinson JM (2013) Trophoblast cell fusion and differentiation are mediated by both the protein kinase C and a pathways. PLoS One 8: doi: 10.1371/journal.pone.0081003. [View Article] [PubMed] [Google Scholar]
- Desforges M, Mynett KJ, Jones RL, Greenwood SL, Westwood M, et al. (2008) The SNAT4 isoform of the system A amino acid transporter is functional in human placental microvillous plasma membrane. J Physiol 587: 61-72. doi: 10.1113/jphysiol.2008.161331. [View Article] [PubMed] [Google Scholar]
- Jones HN, Powell TL, Jansson T (2007) Regulation of placental nutrient transport--a review. Placenta 2007: 763-74. doi: 10.1016/j.placenta.2007.05.002. [View Article] [PubMed] [Google Scholar]
- Hamilton S, Oomomian Y, Stephen G, Shynlova O, Tower CL, et al. (2012) Macrophages infiltrate the human and rat decidua during term and preterm labor: evidence that decidual inflammation precedes labor. Biol Reprod 86: 39. doi: 10.1095/biolreprod.111.095505. [View Article] [PubMed] [Google Scholar]
- Hamilton S, Oomomian Y, Stephen G, Shynlova O, Tower CL, et al. (2012) Macrophages Infiltrate the Human and Rat Decidua During Term and Preterm Labor: Evidence That Decidual Inflammation Precedes Labor. Biol Reprod. Biol Reprod 2012: 1-9. doi: 10.1095/biolreprod.111.095505. [View Article] [PubMed] [Google Scholar]
- Kim JS, Romero R, Kim MR, Kim YM, Friel L, et al. (2008) Involvement of Hofbauer cells and maternal T cells in villitis of unknown aetiology. Histopathology 2008: 457-64. doi: 10.1111/j.1365-2559.2008.02964.x. [View Article] [PubMed] [Google Scholar]